Нейро́н или нервная клетка (от др.-греч. νεῦρον «волокно; нерв») — узкоспециализированная клетка. Нейрон — электрически возбудимая клетка, которая предназначена для приёма извне, обработки, хранения, передачи и вывода вовне информации с помощью электрических и химических сигналов.

Типичный нейрон состоит из тела клетки, дендритов и одного аксона. Нейроны могут соединяться один с другим, формируя нервные сети. По отношению к границе нервной системы и направлению передачи информации нейроны разделяют на рецепторные (граничные, получают сигналы извне, формируют на их основании и передают информацию в нервную систему), эффекторные (граничные, передают сигналы из нервной системы во внешние клетки) и вставочные (внутренние для нервной системы).
Сложность и многообразие функций нервной системы определяется взаимодействием между нейронами, а также между нейронами и мышцами и железами. Это взаимодействие обеспечивается набором различных сигналов, передаваемых с помощью ионов. Ионы генерируют электрический заряд (потенциал действия), который движется по телу нейрона.
Впервые нейроны были обнаружены в 1837 году Яном Пуркине при изучении клеток мозжечка[1].
Важное значение для науки имело изобретение метода Гольджи в 1873 году, позволявшего окрашивать отдельные нейроны[2][3]. Термин «нейрон» (нем. Neuron) для обозначения нервных клеток введён Г. В. Вальдейером в 1891 году[4][5].
Строение нейронов
править
Тело клетки
правитьВ статье не хватает ссылок на источники (см. рекомендации по поиску). |
Тело нервной клетки состоит из протоплазмы (цитоплазмы и ядра), ограниченной снаружи клеточной мембраной (плазмалемма, называемая также — у нейронов — нейролеммой) из липидного бислоя. Липиды состоят из гидрофильных головок и гидрофобных хвостов. Липиды располагаются гидрофобными хвостами друг к другу, образуя гидрофобный слой из головок. Этот слой пропускает только жирорастворимые вещества (напр. кислород и углекислый газ). На мембране находятся белки: в форме глобул на поверхности, на которых можно наблюдать наросты полисахаридов (гликокаликс), благодаря которым клетка воспринимает внешнее раздражение, и интегральные белки, пронизывающие мембрану насквозь, в которых находятся ионные каналы.
Тело нейрона имеет диаметр от 3 до 130 мкм. Тело содержит ядро (с большим количеством ядерных пор) и органеллы (в том числе сильно развитый шероховатый ЭПР с активными рибосомами, аппарат Гольджи), а также отростки. Выделяют два вида отростков: дендриты и аксон. Нейрон имеет развитый цитоскелет, который проникает в его отростки. Цитоскелет поддерживает форму клетки, его нити служат «рельсами» для транспорта органелл и упакованных в мембранные пузырьки веществ (например, нейромедиаторов). Цитоскелет нейрона состоит из фибрилл разного диаметра: микротрубочки (Д = 20—30 нм) состоят из белка тубулина и тянутся от нейрона по аксону вплоть до нервных окончаний. Нейрофиламенты (Д = 10 нм) вместе с микротрубочками обеспечивают внутриклеточный транспорт веществ. Микрофиламенты (Д = 5 нм) состоят из белков актина и, в отличие от других клеток, не содержат миозина, что делает невозможным сокращение в этих клетках, сами микрофиламенты особенно выражены в растущих нервных отростках и в нейроглии. Нейроглия, или просто глия (от др.-греч. νεῦρον — волокно, нерв + γλία — клей) — совокупность вспомогательных клеток нервной ткани. Составляет около 40 % объёма ЦНС. Количество глиальных клеток в мозге примерно равно количеству нейронов.
В теле нейрона выявляется развитый синаптический аппарат, гранулярная эндоплазматическая сеть нейрона окрашивается базофильно и известна под названием «тигроид». Тигроид проникает в начальные отделы дендритов, но располагается на заметном расстоянии от начала аксона, что служит гистологическим признаком аксона. Нейроны различаются по форме, числу отростков и функциям. В зависимости от функции выделяют чувствительные, эффекторные (двигательные, секреторные) и вставочные. Чувствительные нейроны воспринимают раздражения, преобразуют их в нервные импульсы и передают в мозг. Эффекторные (от лат. effectus — действие) — вырабатывают и посылают команды к рабочим органам. Вставочные — осуществляют связь между чувствительными и двигательными нейронами, участвуют в обработке информации и выработке команд.
Различается антероградный (от тела) и ретроградный (к телу) аксонный транспорт, осуществляемые с помощью кинезин-динеинового механизма (кинезин отвечает за антероградный ток, динеин — за ретроградный).
Дендриты и аксон
правитьВ статье не хватает ссылок на источники (см. рекомендации по поиску). |

Аксон — длинный отросток нейрона. Приспособлен для проведения возбуждения и информации от тела нейрона (нейросомы) к другому нейрону (иногда к этому же, см. нейронные ловушки), или же от нейрона к исполнительному органу. Дендриты — короткие и сильно разветвлённые отростки нейрона, служащие главным местом для образования влияющих на нейрон возбуждающих и тормозных синапсов (разные нейроны имеют различное соотношение длины аксона и дендритов), и которые передают возбуждение к телу нейрона. Нейрон может иметь несколько дендритов и обычно только один аксон. Один нейрон может иметь связи со многими (до 20 тысяч) другими нейронами.
Дендриты делятся дихотомически, аксоны же дают коллатерали. В узлах ветвления обычно сосредоточены митохондрии.
Дендриты не имеют миелиновой оболочки, аксоны же могут её иметь. Местом генерации потенциала действия (ПД, «спайк») у большинства нейронов является аксонный холмик (триггерная зона нейрона) — образование в месте отхождения аксона от нейросомы. Мембранный потенциал покоя в этом месте немного меньше, то есть порог деполяризации тоже меньше. Также здесь большое количество кальциевых и натриевых каналов, которые участвуют в генерации импульса.
Синапс
правитьВ статье не хватает ссылок на источники (см. рекомендации по поиску). |
Си́напс (греч. σύναψις, от συνάπτειν — обнимать, обхватывать, пожимать руку) — специализированное образование, которое обеспечивает контакт между двумя нейронами или между нейроном и получающей сигнал эффекторной клеткой (клетки возбудимых тканей). Служит для передачи нервного импульса между двумя клетками, причём в ходе синаптической передачи амплитуда и частота сигнала могут регулироваться. Одни синапсы вызывают деполяризацию нейрона и являются возбуждающими, другие — гиперполяризацию и являются тормозными. Обычно для возбуждения нейрона необходимо раздражение от нескольких возбуждающих синапсов.
Термин был введён английским физиологом Чарльзом Шеррингтоном в 1897 г[6].

Механизм создания и проведения потенциала действия
правитьВ статье не хватает ссылок на источники (см. рекомендации по поиску). |
В 1937 году Джон Захари Младший определил, что гигантский аксон кальмара может быть использован для изучения электрических свойств аксонов. Аксоны кальмара были выбраны из-за того, что они намного крупнее человеческих. Если вставить внутрь аксона электрод, то можно замерить его мембранный потенциал.
Мембрана аксона содержит в себе потенциал-зависимые ионные каналы. Они позволяют аксону генерировать и проводить по своему телу электрические сигналы, называемые потенциалами действия. Эти сигналы образуются и распространяются благодаря электрически заряженным ионам натрия (Na+), калия (K+), хлора (Cl-), кальция (Ca2+).
Давление, растяжение, химические факторы или изменение мембранного потенциала могут активировать нейрон. Происходит это вследствие открытия ионных каналов, которые позволяют ионам пересекать мембрану клетки и соответственно изменять мембранный потенциал.
Тонкие аксоны расходуют меньше энергии и метаболических веществ для проведения потенциала действия, но толстые аксоны позволяют проводить его быстрее.
Для того, чтобы проводить потенциалы действия более быстро и менее энергозатратно, нейроны могут использовать для покрытия аксонов специальные глиальные клетки, называемые олигодендроцитами в ЦНС, или шванновскими клетками в периферической нервной системе. Эти клетки покрывают аксоны не полностью, оставляя промежутки на аксонах, открытые внеклеточному веществу. В этих промежутках повышенна плотность ионных каналов. Они называются перехватами Ранвье. Через них и проходит потенциал действия посредством электрического поля между промежутками.
Классификация
правитьВ статье не хватает ссылок на источники (см. рекомендации по поиску). |
Структурная классификация
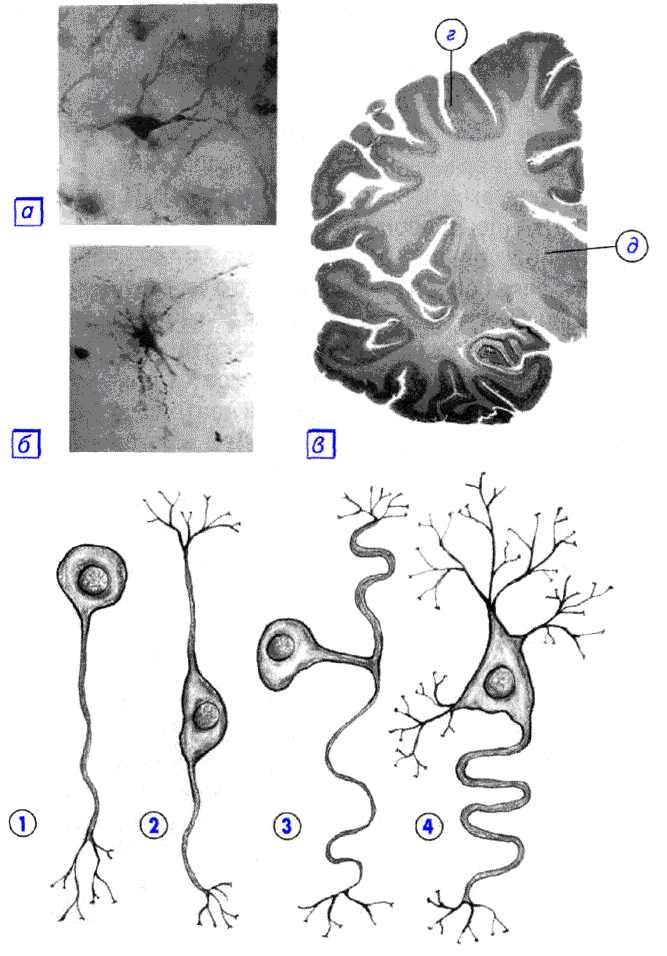
правитьНа основании числа и расположения дендритов и аксона нейроны делятся на безаксонные, униполярные нейроны, псевдоуниполярные нейроны, биполярные нейроны и мультиполярные (много дендритных стволов, обычно эфферентные) нейроны[7].
Безаксонные нейроны — небольшие клетки, сгруппированы вблизи спинного мозга в межпозвоночных ганглиях, не имеющие анатомических признаков разделения отростков на дендриты и аксоны. Все отростки у клетки очень похожи. Функциональное назначение безаксонных нейронов слабо изучено.
Униполярные нейроны — нейроны с одним отростком, присутствуют, например в сенсорном ядре тройничного нерва в среднем мозге. Многие морфологи считают, что униполярные нейроны в теле человека и высших позвоночных не встречаются.
Биполярные нейроны — нейроны, имеющие один аксон и один дендрит, расположенные в специализированных сенсорных органах — сетчатке глаза, обонятельном эпителии и луковице, слуховом и вестибулярном ганглиях.
Мультиполярные нейроны — нейроны с одним аксоном и несколькими дендритами. Данный вид нервных клеток преобладает в центральной нервной системе.
Псевдоуниполярные нейроны — являются уникальными в своём роде. От тела отходит один отросток, который сразу же Т-образно делится. Весь этот единый тракт покрыт миелиновой оболочкой и структурно представляет собой аксон, хотя по одной из ветвей возбуждение идёт не от, а к телу нейрона. Структурно дендритами являются разветвления на конце этого (периферического) отростка. Триггерной зоной является начало этого разветвления (то есть находится вне тела клетки). Такие нейроны встречаются в спинальных ганглиях.
Функциональная классификация
правитьПо положению в рефлекторной дуге различают афферентные нейроны (чувствительные нейроны), эфферентные нейроны (часть из них называется двигательными нейронами, иногда это не очень точное название распространяется на всю группу эфферентов) и интернейроны (вставочные нейроны).
Афферентные нейроны (чувствительный, сенсорный, рецепторный, или центростремительный). К нейронам данного типа относятся первичные клетки органов чувств и псевдоуниполярные клетки, у которых дендриты имеют свободные окончания.
Эфферентные нейроны (эффекторный, двигательный, моторный, или центробежный). К нейронам данного типа относятся конечные нейроны — ультиматные и предпоследние — не ультиматные.
Ассоциативные нейроны (вставочные, или интернейроны) — группа нейронов осуществляет связь между эфферентными и афферентными.
Секреторные нейроны — нейроны, секретирующие высокоактивные вещества (нейрогормоны). У них хорошо развит комплекс Гольджи, аксон заканчивается аксовазальными синапсами.
Морфологическая классификация
правитьМорфологическое строение нейронов многообразно. При классификации нейронов применяют несколько принципов:
- учитывают размеры и форму тела нейрона;
- количество и характер ветвления отростков;
- длину аксона и наличие специализированных оболочек.
По форме клетки, нейроны могут быть сферическими, зернистыми, звездчатыми, пирамидными, грушевидными, веретеновидными, неправильными и т. д. Размер тела нейрона варьирует от 5 мкм у малых зернистых клеток до 120—150 мкм у гигантских пирамидных нейронов.
По количеству отростков выделяют следующие морфологические типы нейронов[8]:
- униполярные (с одним отростком) нейроциты, присутствующие, например, в сенсорном ядре тройничного нерва в среднем мозге;
- псевдоуниполярные клетки, сгруппированные вблизи спинного мозга в межпозвоночных ганглиях;
- биполярные нейроны (имеют один аксон и один дендрит), расположенные в специализированных сенсорных органах — сетчатке глаза, обонятельном эпителии и луковице, слуховом и вестибулярном ганглиях;
- мультиполярные нейроны (имеют один аксон и несколько дендритов), преобладающие в ЦНС.
Также нейроны классифицируются по воздействию (тормозные и возбуждающие) и секретируемому медиатору (ацетилхолин, ГАМК и т. д., последних ещё в 1985 году было известно более 50, и ещё тогда исследователями выделялись тысячи типов клеток.[9]
Развитие и рост нейрона
правитьВ статье не хватает ссылок на источники (см. рекомендации по поиску). |

Вопрос о делении нейронов в настоящее время остаётся дискуссионным. По одной из версий, нейрон развивается из небольшой клетки-предшественницы, которая перестаёт делиться ещё до того, как выпустит свои отростки. Первым начинает расти аксон, а дендриты образуются позже. На конце развивающегося отростка нервной клетки появляется утолщение, которое прокладывает путь через окружающую ткань. Это утолщение называется конусом роста нервной клетки. Он состоит из уплощённой части отростка нервной клетки со множеством тонких шипиков. Микрошипики имеют толщину от 0,1 до 0,2 мкм и могут достигать 50 мкм в длину, широкая и плоская область конуса роста имеет ширину и длину около 5 мкм, хотя форма её может изменяться. Промежутки между микрошипиками конуса роста покрыты складчатой мембраной. Микрошипики находятся в постоянном движении — некоторые втягиваются в конус роста, другие удлиняются, отклоняются в разные стороны, прикасаются к субстрату и могут прилипать к нему.
Конус роста заполнен мелкими, иногда соединёнными друг с другом, мембранными пузырьками неправильной формы. Под складчатыми участками мембраны и в шипиках находится плотная масса перепутанных актиновых филаментов. Конус роста содержит также митохондрии, микротрубочки и нейрофиламенты, аналогичные имеющимся в теле нейрона.
Микротрубочки и нейрофиламенты удлиняются главным образом за счёт добавления вновь синтезированных субъединиц у основания отростка нейрона. Они продвигаются со скоростью около миллиметра в сутки, что соответствует скорости медленного аксонного транспорта в зрелом нейроне. Поскольку примерно такова и средняя скорость продвижения конуса роста, возможно, во время роста отростка нейрона в его дальнем конце не происходит ни сборки, ни разрушения микротрубочек и нейрофиламентов. Новый мембранный материал добавляется у окончания. Конус роста — это область быстрого экзоцитоза и эндоцитоза, о чём свидетельствует множество находящихся здесь пузырьков. Мелкие мембранные пузырьки переносятся по отростку нейрона от тела клетки к конусу роста с потоком быстрого аксонного транспорта. Мембранный материал синтезируется в теле нейрона, переносится к конусу роста в виде пузырьков и включается здесь в плазматическую мембрану путём экзоцитоза, удлиняя таким образом отросток нервной клетки.
Росту аксонов и дендритов обычно предшествует фаза миграции нейронов, когда незрелые нейроны расселяются и находят себе постоянное место.
При этом у нейронов распространён эндомитоз, приводящий к соматической полиплоидии[10][11].
Свойства и функции нейронов
правитьВ статье не хватает ссылок на источники (см. рекомендации по поиску). |
- Свойства
- Наличие трансмембранной разницы потенциалов (до 90 мВ), наружная поверхность электроположительна по отношению к внутренней поверхности.
- Очень высокая чувствительность к некоторым химическим веществам и электрическому току (вообще говоря, чувствительность к самым разнообразным химическим соединениям, нарушениям биохимического гомеостаза и т. д.).
- Электровозбудимость вышеупомянутыми, и её обратная сторона — затормаживаемость.
- Порог возбуждения для раздражителей.
- Способность к нейросекреции, то есть к синтезу и выделению особых веществ (нейромедиаторов), в окружающую среду или синаптическую щель.
- Высокое энергопотребление, высокий уровень энергетических процессов, что обуславливает необходимость постоянного притока основных источников энергии — глюкозы и кислорода, необходимых для окисления.
- Усиление сигнала.
- Переходная характеристика.
- Перекодирование (преобразование ритма).
- Инерционность.
- Обучаемость (в основном нейроны экранных центров).
- Адаптируемость (в том числе к ритму — усвоение ритма).
- Модулируемость (ретикулярной формацией, электротоном, гормонами).
- Спонтанная активность определённого ритма и уровня (в том числе весьма близкого к нулю).
- Нейроантигенность[12].
- Выделение всеми терминалями аксона данного нейрона 1 и того же: медиатора, или «коктейля» медиаторов (уточнённый принцип Дейла[13]).
- Функции
- Приёмная функция. Синапсы — точки контакта, от рецепторов и нейронов получаем информацию в виде последовательностей нервных импульсов («спайков»).
- Интегративная функция. В результате обработки информации на выходе нейрона формируется сигнал, несущий информацию, выработанную из всех входных сигналов.
- Проводниковая функция. От нейрона по аксону идёт информация в виде потенциалов действия к синапсам.
- Передающая функция. Нервный импульс, достигнув окончания аксона, которое уже входит в структуру синапса, обуславливает выделение медиатора — непосредственного передатчика возбуждения к другому нейрону или исполнительному органу.
- Значение
По мнению известного американского нейрофизиолога Майкла Грациано, характер связей между нейронами того или иного мозга определяет суть данного мозга и его отличие от прочих[14].
См. также
правитьПримечания
править- ↑ Нейроперсоналии: Ян Пуркинье, первооткрыватель клеток мозжечка. Дата обращения: 6 марта 2021. Архивировано 3 января 2022 года.
- ↑ Camillo Golgi. Sulla struttura della sostanza grigia del cervelo (неопр.) // Gazzetta Medica Italiana. Lombardia. — 1873. — Т. 33. — С. 244—246. Архивировано 27 августа 2016 года.
- ↑ Айзек Азимов. Краткая история биологии. — Рипол Классик, 2013. — С. 114.
- ↑ Jean-Pierre Changeux,Laurence Garey. Neuronal Man – The Biology of Mind. — Princeton University Press, 1997. — С. 28.
- ↑ А именно, термин «нейрон» встречается в цитате «Das Nervensystem besteht aus zahlreichen, untereinander anatomisch wie genetisch nicht zusammenhängenden Nerveneinheiten (Neuronen)» из следующего источника:Heinrich Wilhelm Gottfried von Waldeyer-Hartz. Ueber einige neuere Forschungen im Gebiete der Anatomie des Centralnervensystems (нем.) // Deutsche medicinische Wochenschrift : magazin. — 1891. — Bd. 17, Nr. 50. — S. 1352. — doi:10.1055/s-0029-1206907.
- ↑ С. А. Осиповский; H. Н. Боголепов. Синапс // Большая медицинская энциклопедия / под ред. Б. В. Петровского. — 3-е изд. — Т. 23. Архивировано 24 августа 2017 года.
- ↑ И. В. Гайваронский. Анатомия центральной нервной системы. Краткий курс / Г. И. Ничипорук. — 4-е изд., перераб. и доп. — СПб.: ЭЛБИ-СПб, 2010. — С. 3—4. — 108 с.
- ↑ см. рис. Архивная копия от 20 июля 2014 на Wayback Machine
- ↑ Arnold J. Mandell, From Molecular Biological Simplification to More Realistic Central Nervous System Dynamics: An Opinion, in Psychiatry: Psychobiological Foundations of Clinical Psychiatry 3:2, J. O. Cavenar, et al., eds. (New York: Lippincott, 1985). cited by: Gleick, James. Chaos: Making a New Science. Penguin books, New York NY, 1987. Page 299. URL: [1] Архивная копия от 8 ноября 2020 на Wayback Machine
- ↑ Кирсанова, Ирина Александровна. «Соматическая полиплоидия в центральной нервной системе лёгочных брюхоногих моллюсков.» (2003).
- ↑ Ю. А. Современные аспекты соматической полиплоидии (О первом Всесоюзном симпозиуме по соматической полиплоидии у животных, Ереван, 10-12 ноября 1971 г.) //Հայաստանի կենսաբանական հանդես Biological Journal of Armenia Биологический журнал Армении. — 1972. — Т. 25. — №. 1. — С. 112—115.
- ↑ Ашмарин, Игорь Петрович. Загадки и откровения биохимии памяти. — Издательство Ленингр. университета, 1975.
- ↑ Osborne, N. N. Dale’s Principle and Communication Between Neurones / ed. N. N. Osborne // England: Pergamon Press, 1983. — 204 p. — Режим доступа: DOI: 10.1016/C2013-0-03680-8
- ↑ Грациано, 2021, Глава 8.
{kind=link}
{kind=link}
Литература
править- Поляков Г. И. О принципах нейронной организации мозга, М: МГУ, 1965
- Косицын Н. С. Микроструктура дендритов и аксодендритических связей в центральной нервной системе. М.: Наука, 1976, 197 с.
- Немечек С. и др. Введение в нейробиологию, Avicennum: Прага, 1978, 400 c.
- Блум Ф., Лейзерсон А., Хофстедтер Л. Мозг, разум и поведение
- Мозг (сборник статей: Д. Хьюбел, Ч. Стивенс, Э. Кэндел и дp. — выпуск журнала Scientific American (сентябрь 1979)). М. :Миp, 1980
- Савельева-Новосёлова Н. А., Савельев А. В. Устройство для моделирования нейрона. А. с. № 1436720, 1988
- Савельев А. В. Источники вариаций динамических свойств нервной системы на синаптическом уровне // журнал “Искусственный интеллект”, НАН Украины. — Донецк, Украина, 2006. — № 4. — С. 323—338.
- Майкл Грациано. Наука сознания. Современная теория субъективного опыта = Michael S. A. Graziano. Rethinking Consciousness: A Scientific Theory of Subjective Experience. — М.: Альпина нон-фикшн, 2021. — 254 с. — (Книги Политеха). — ISBN 978-5-00139-208-8.
Ссылки
править- Нейромодель RF-PSTH (симулирующая структуру рецептивного поля (РП) и выходной нейронный сигнал PSTH)
- Научно-популярный фильм «Нервная клетка» на YouTube
- Что такое нейрон? | Перевод научной статьи Dr. C. George Boeree
В статье не хватает ссылок на источники (см. рекомендации по поиску). |