Листосте́бельные мхи, или бри́евые мхи, или брио́псиды (лат. Bryópsida) — класс мхов. В отличие от других моховидных, тело гаметофита листостебельных мхов расчленено на стебель и листья.
| Листостебельные мхи | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | ||||||||||||
| Научная классификация | ||||||||||||
|
Домен: Царство: Подцарство: Надотдел: Отдел: Класс: Листостебельные мхи |
||||||||||||
| Международное научное название | ||||||||||||
| Bryopsida (Limpr.) Rothm. | ||||||||||||
| Подклассы | ||||||||||||
|
См. текст |
||||||||||||
| ||||||||||||
Класс листостебельные мхи является крупнейшим классом мхов, в котором содержится 95 % всех видов. Он включает примерно 14 500 видов, растущих по всему миру.
Строение
правитьСтебель
правитьСтебель (каулидий) у листостебельных мхов имеется всегда, покрыт листьями. Органы размножения располагаются сверху (акрокарпные или верхоплодные мхи) или по бокам (плеврокарпные или бокоплодные мхи) стебля. Акрокарпные мхи имеют прямостоящий стебель (ортотропные мхи), плеврокарпные — горизонтальный стебель (плагиотропные мхи), лежащий, свисающий или плавающий. На самом деле у плагиотропных мхов органы размножения сидят на верхушках укороченных боковых побегов, так что разделение на акрокарпные и плеврокарпные мхи носит формальный характер и допускает множество переходных форм.
В поперечном разрезе стебель округлый, иногда овальный, угловатый или ребристый. Может быть из однородных клеток, но у большинства мхов состоит из дифференцированных. В последнем случае образуется кора или склеродерма, из механической ткани, внутри которой находится основная, проводящая, ткань. Отмершие клетки внутренней коры образуют снаружи эпидермис, один или несколько слоев омертвевших клеток без живого содержимого, служащих для сохранения воды.
Механическую ткань образуют стереиды — вытянутые в длину, узкие, прозенхимные клетки. Обычно имеют желтоватый, бурый, красно-бурый, пурпурный или почти чёрный цвет. Стенки стереид толстые, иногда до почти полного исчезновения просвета. Во внутренних частях стенок имеются водопроводящие поры. Внутренняя поверхность стенки может образовывать папиллообразные выросты, густой сетью прорастающие в полость клетки. Они служат для запасения воды, благодаря чему клетки могут набухать. Основная функция внутренней коры — обеспечивать прочность стебля.
У некоторых мхов наружный слой коры образуют клетки гиалодермы — без хлоропластов, с тонкими, прозрачными стенками и широким просветом.
Устьица в эпидерме стебля отсутствуют у всех листостебельных мхов. Это связано с тем, что они, по сути, и не нужны мхам, с их отсутствием настоящей покровной ткани, требующей регулировки процессов поглощения и выделения. Как поглощение воды, так и её испарение осуществляются сразу всей поверхностью гаметофита.
Чем глубже, тем менее толстые стенки у клеток коры, которая постепенно переходит в основную ткань. Реже кора резко отграничена от основной ткани (Meesia longiseta). Основная ткань состоит из однородных паренхимных клеток, обильно заполненных цитоплазмой, хлоропластами, крахмалом, жирными маслами. В поперечном разрезе клетки часто колленхиматически (в форме треугольных вздутий на углах) утолщены. Основная ткань обеспечивает фотосинтез, служит для сохранения воды, запасов крахмала и масла, а также продуктов секреции (оксалата кальция). Главная функция основной ткани — водопроводящая.
Иногда в основной ткани могут находиться группы мелких тонкостенных клеток, продолжающие в стебле жилки листьев, листовые следы. Одни заканчиваются в основной ткани стебля и не доходят до центрального пучка (так называемые ложные листовые следы), другие — соединяются с центральным пучком (настоящие листовые следы). Настоящие листовые следы могут быть дифференцированы на ткани (сплахнум).
Центральный пучок, или проводящий пучок проходит по оси стебля, состоит из вытянутых в длину, с косыми поперечными перегородками, большей частью тонкостенных клеток с узким просветом. Иногда клетки центрального пучка толстостенные и окрашенные (Dicranum scoparium, виды Hypnum). Обычно проводящий пучок четко отграничен от основной ткани. Центральный пучок обеспечивает проведение воды и её сохранение. Имеется не у всех видов и родов. В случае отсутствия центрального пучка его функции выполняет основная ткань, заполняющая стебель целиком.
-
 Поперечный разрез стебля Rhizomnium punctatum
Поперечный разрез стебля Rhizomnium punctatum -
 Он же в большем увеличении
Он же в большем увеличении -
 Orthotrichum lyellii
Orthotrichum lyellii -
 Hedwigia ciliata
Hedwigia ciliata -
 Catoscopium nigritum
Catoscopium nigritum -
 Anomobryum concinnatum
Anomobryum concinnatum
_6649.JPG)
_6652.JPG)
_1919.JPG)
_3090.JPG)
_7667.JPG)
_2522.JPG)
Из любой поверхностной клетки стебля могут образовываться волосовидные образования — ризоиды. Это однорядные многоклеточные нити с бурыми, фиолетовыми или красноватыми продольными оболочками и с косыми поперечными перегородками. С помощью ризоидов мхи крепятся к субстрату и адсорбируют воду из окружающей среды. Кроме того, ризоиды способны к фотосинтезу. Формируются по всей поверхности стебля, но чаще всего — у основания прямостоячего стебля или на брюшной, обращённой к субстрату, стороне ползучего или лежачего стебля. Иногда ризоиды покрывают весь стебель густым войлоком, светлым или окрашенным в бурый, фиолетовый или красноватый цвет. У некоторых мхов ризоиды скручиваются в длинные тяжи.
Ризоидный войлок поднимает воду из почвы и долго сохраняет её в капиллярных пространствах между ризоидами и растением. Ризоиды водных мхов прикрепляют их к субстрату, адсорбирующая функция для них не столь актуальна. Для более надёжного крепления на концах ризоидов водных мхов образуются вильчатые разветвления, иногда сплетающиеся в подушечки. С изменением питания и освещения ризоиды могут превратиться в зелёную вторичную протонему.
На верхушке стебля, в пазухах листьев, в месте отхождения листьев от стебля при росте возникают нитевидные образования из нескольких клеток, — булавовидные волоски. Их оконечная клетка выделяет слизь, защищающую точки роста стебля от высыхания.
У некоторых бокоплодных мхов на стебле развиваются выросты стебля — парафиллии (греч. para — возле, греч. phyllon — лист). Они зелёные, нитевидные или листовидные, простые или разветвлённые. Иногда парафиллии вырастают так густо, что окутывают стебель густым зелёным войлоком. Формы парафиллии принимают самые разные, на стебле располагаются безо всякого порядка. Жилок никогда не имеют. Могут не только проводить и удерживать воду, но и выполнять функцию фотосинтеза. Нитевидные парафиллии крепятся к стеблю одной, а листовидные — двумя или более клетками основания.
Стебель никогда не ветвится надвое. Всегда боковые побеги образуются на главном, ниже листа, никогда — в пазухах.
Ветвление мхов бывает двух основных видов: симподиальное и моноподиальное.
Акрокарпные мхи ветвятся в основном по симпоидальному типу. Ветвление слабое, обычно только в верхней части. Инновации, то есть новые боковые ветви, образуются под органами размножения и растут в направлении главного стебля, на верхушке они могут снова давать гаметангии.
Ветвление может идти по симпоидальному типу (побеги возникают по одному), по вильчатому типу (парные побеги) или по пучковидному типу (сразу несколько побегов). Вильчатое и пучковидное ветвление здесь — частный случай симпоидального, где инноваций возникает сразу две или несколько. После отмирания главного стебля инновации, укореняясь, становятся самостоятельными растениями.
У бокоплодных мхов стебли ветвятся моноподиально. Есть явно выраженный главный побег, от которого отходят боковые ветви. На каждой стороне стебля ветви появляются в восходящей (акропетальной) последовательности. Чем моложе побег, тем он ближе к верхушке. Новые ветви отрастают слабо, поскольку главный побег долго не отмирает и продолжает расти.
При правильном чередовании зачатков ветвей ветвление стебля перистое; при повторном ветвлении (образовании на боковых побегах ветвей второго и третьего порядка) — дважды или триждыперистое ветвление.
Рост побегов и развитие ветвей и у верхоплодных, и у бокоплодных мхов зависит от точки роста главного стебля. Если удалить верхушку стебля, то разовьются в побеги покоящиеся почки.
В разделе не хватает ссылок на источники (см. рекомендации по поиску). |
Листья
правитьЛистья (филлидии или филлоиды) всегда сидячие, бесчерешковые, как правило, поперечно прикреплённые к стеблю, расположены всегда по спирали, в три или пять рядов, изредка двурядно. Никогда не располагаются супротивно или мутовчато. Закладываются в восходящей последовательности. Листья образуются делением двусторонней верхушечной клетки, от которой с двух сторон отделяются сегменты. Сегменты делятся перегородками всегда только в одной плоскости. С прекращением верхушечного роста листа его окончательный размер достигается вставочным ростом в его основании.
По расположению на стебле делятся на низовые, срединные и покровные.
Стеблевые листья — срединные, расположены в средней и верхней частях стебля. Веточные листья у большинства мхов не отличаются от стеблевых формой, только более мелкими размерами. Но у некоторых мхов листья на ветках могут принимать иную форму, чем на основном побеге. Расположены очень близко друг к другу, закрывая всю облиственную поверхность стебля.
Покровные листья окружают органы размножения, обычно значительно больше стеблевых, отличаются от них формой, а часто и строением, иногда окраской. Покровные листья, окружающие архегонии, называются перихециальными, а покровные листья, окружающие антеридии, — перигониальными.
Низовые листья сильно редуцированные, намного меньше стеблевых, чешуевидные. Развиваются в нижней надземной или подземной части стебля у мхов, имеющих ризомы. Часто не имеют хлорофилла.
Листья мхов простые, цельные, по краю могут быть зубчатыми и крайне редко глубоко разделёнными. Листовая пластинка чаще всего однослойная, реже она целиком или частично двуслойная или многослойная. Пластинка может быть плоской, чашевидной, ложковидной, желобчатой, складчатой, килеватой, бороздчатой или волнистой. Поверхность может быть гладкой и блестящей, или бархатистой и тусклой. Клетки листа богаты хлоропластами и выполняют функцию фотосинтеза. Жилка, если она вообще развита, проходит посередине листа и состоит из толстостенных, вытянутых клеток. Кроме механического удержания листа, жилка обеспечивает проведение пластических веществ и воды. Кожицы, устьиц на листе никогда не бывает.
В пластинке листа различают основание, верхушку, край, кайму, крылья и ушки.
Основанием называется место прикрепления листа к стеблю. Противоположный конец называют верхушкой. Листовые крылья или листовые ушки развиваются на углах основания листа. Ушки резко отличаются формой, окраской и размерами от остальных клеток и служат для сохранения воды. Кайму листа образуют длинные и узкие клетки, расположенные в один или несколько рядов по краю листа.
-
 Основание Bryum alpinum
Основание Bryum alpinum -
 Верхушка Bryum capillare
Верхушка Bryum capillare -
 Край Mnium spinosum
Край Mnium spinosum -
 Крылья Scleropodium purum
Крылья Scleropodium purum
_0197.JPG)
_8411.JPG)
_9424.JPG)

По форме листья очень разнообразны — от округлых до шиловидных. Наиболее часто встречаются формы:
- округлая;
- яйцевидная;
- эллиптическая;
- языковидная;
- ланцетная;
- шиловидная.
Строение краёв листьев разнообразно. Отворачиваясь, загибаясь или отгибаясь, край листа создаёт микроскопические полости, где удерживается капиллярная вода. Завернутый край листа характерен для мха, обитающего в условиях периодической засухи, поскольку выполняет защитную функцию, предохраняя лист от иссушения.
Клетки листа бывают двух основных типов: паренхиматические — обычно округлые или многоугольные (часто квадратные и шестиугольные), почти одинаковой длины и ширины, и прозенхиматические — узкие, вытянутые в длину, прямые, извитые, червеобразные с заострёнными, заходящими друг за друга концами. Клетки пластинки листа редко одинаковые. Верхняя часть листа обычно состоит из толстостенных паренхиматических мелких клеток, иногда мамиллозных (конусообразные выросты клеток без утолщения клеточных оболочек) или папиллозных. Клетки основания всегда несколько крупнее и резко отличаются от верхних клеток по форме.
Папиллы (утолщения клеточной оболочки) наиболее характерны для бриевых мхов. Они сильно увеличивают всасывающую поверхность клетки, способствуют более быстрому забору воды и проведению её внутрь клетки. Особенно часто развиваются папиллы на клетках мхов, растущих в сухих и сильно освещённых местах, но широко распространены и у произрастающих в условиях избыточной влажности.
У ряда систематически далёких групп мхов проявляется чёткое обособление ассимиляционных и водоносных элементов листа.
У верхоплодных мхов жилка листа обычно простая, неразветвлённая. Бокоплодные мхи могут иметь совершенно различные жилки или не иметь их вовсе даже у представителей одного семейства. Жилка может быть простая — в одну полосу; двойная, расходящаяся сразу же от основания листа на два луча; вильчатая — простая жилка с короткими ответвлениями.
В большинстве случаев жилка проходит лишь по нижней стороне листа, и лишь у некоторых мхов она есть и на верхней стороне. На нижней стороне жилки часто вырастают разнообразные структурные образования. В семействе поттиевых у родов алоина (Aloina Kindb.) и кроссидиум (Crossidium Jur.) в верхней части листа от жилки прорастают многоклеточные, иногда разветвлённые нити, богатые хлоропластами, которые часто сплетаются в подушку.
Ассимиляционные пластиночки и нити на листьях мхов выполняют, вероятно, одни и те же функции: у одних мхов ассимиляционную, у других — обеспечивающую всасывание воды. При подсыхании большинства таких мхов края листьев заворачиваются, прикрывая плотно сверху пластиночки и нити. Обламывающиеся нити листа способны к вегетативному размножению.
У большинства верхоплодных мхов жилка имеет сложное строение. Она состоит из указателей, сопроводителей, стереидных пучков и наружных клеток. Паренхиматические клетки жилки листа с широким просветом, тонкостенные, бедные плазматическим содержимым, расположенные в один, реже в два ряда, называют указателями. Они проводят воду и на продольных стенках нередко имеют поры. В сырую погоду указатели заполнены водой. Вследствие этого жилка разбухает и вытягивается, а вместе с ней растягивается пластинка листа. В сухую погоду жилка подсыхает, теряет натяжение, сгибается внутрь, а с нею изгибается и пластинка листа. Сопроводители — тонкостенные, вытянутые в длину клетки малого диаметра, объединённые в тяж и внешне напоминающие клетки центрального пучка стебля. Они почти всегда находятся на верхней стороне указателей. Сопроводители богаты плазмой. Указатели и сопроводители называются характерными клетками жилки. К ним обычно со спинной стороны или с обеих сторон, реже только с брюшной, прилегают стереиды.
Наружные клетки (обычно с довольно широким просветом) образуют эпидерму жилки. Они обычно лежат только с одной стороны жилки — или спинной, или брюшной. Их функция — защитная. Стереиды жилки — толстостенные, вытянутые в длину и соединённые в тяжи клетки. Они сходны с лубяными волокнами сосудистых растений и по строению не отличаются от стереидных клеток стебля. Механическая ткань жилки образует один лентовидный стереидный пучок в середине жилки, у ряда видов — два, спинной и брюшной, разделённые указателями. Стереиды укрепляют жилку.
-
 Лист Anomobryum concinnatum
Лист Anomobryum concinnatum -
 Лист Catoscopium nigritum
Лист Catoscopium nigritum -
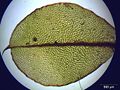 Лист Cinclidium stygium
Лист Cinclidium stygium
_2505.JPG)
_7639.JPG)

В разделе не хватает ссылок на источники (см. рекомендации по поиску). |
Органы размножения
правитьОрганы полового размножения мхов чаще всего собраны в группы. Их обычно окружают покровные листья. У акрокарпных мхов гаметангии образуются на верхушках главных побегов. У плеврокарпных на очень коротких боковых ветвях. Между антеридиями (мужские половые органы) и архегониями (женские половые органы) часто находятся нитевидные или булавовидные парафизы. Парафизы антеридиальныех побегов способны поглощать и удерживать влагу. Парафизы вокруг архегония покрывают его, защищая от неблагоприятных условий.
Одни виды мхов однодомные, другие — двудомные. Двудомность у мхов, видимо, относительна, так как на одной и той же протонеме могут формироваться и женские и мужские экземпляры. В условиях недостаточного питания на протонеме формируются мужские гаметофиты, а при хорошем питании — женские.
Листостебельные мхи обладают ярко выраженным половым диморфизмом. Женские растения, которым необходимо питать молодой спорофит, обычно крупнее и более развиты. Мужские растения у двудомных видов часто сильно редуцированы. Иногда мужские растения просто карлики, отмирающие после образования антеридиев.
У некоторых видов гипнума споры прорастают прямо на листьях женских растений и из них вырастают карликовые мужские растеньица.
У одних видов мхов генеративные и вегетативные побеги дифференцируются. У других видов такой дифференциации нет.
Зрелые антеридии — эллипсоидальные, булавовидные, реже шаровидные (у рода буксбаумия) тела, часто на короткой многоклеточной ножке. Под однослойной стенкой антеридия располагается сперматогенная ткань, из которой формируются жгутиковые сперматозоиды. Внутри перигония, обычно не одновременно, формируется большое число антеридиев. Зрелые антеридии раскрываются при выпадении дождя или росы. Сперматозоиды по воде движутся к архегонию.
Архегоний обычно имеет форму бутыльчатого тела на массивной ножке. Верхняя узкая часть архегония называется шейкой, нижняя расширенная — брюшком. Внутри шейки расположены шейковые канальцевые клетки, в брюшке — одна-две брюшные канальцевые клетки. На дне брюшка находится одна крупная яйцеклетка. Брюшная клетка и канальцевые клетки шейки зрелого архегония покрываются слизью. Покровные клетки шейки, как и у антеридия, выделяют слизь и разрываются, образуя проход, ведущий к яйцеклетке. Сперматозоиды благодаря хемотаксису движутся к этой слизи и оплодотворяют яйцеклетку.
Хотя в перихеции развивается несколько архегониев, у большинства видов оплодотворяется только один из них. Соответственно, вырастает только один спорогон. Однако у некоторых видов в одном перихеции оплодотворяется сразу несколько архегониев из которых вырастает несколько спорогонов.
В результате оплодотворения яйцеклетки в брюшке архегония образуется зигота. Нижняя клетка в процессе деления образует нижнюю часть ножки и стопу, прорастающую сквозь стенку брюшка архегония до тканей гаметофита. Из верхней (апикальной) клетки поделившейся зиготы формируется коробочка.
По мере роста молодого спорофита развивается стеблеобразное тело, которое дифференцируется затем на коробочку и ножку со стопой. Удлиняющийся спорофит разрывает в поперечнике окружающий его разросшийся архегоний. Коробочка приподнимает верхнюю часть разорванного архегония в виде колпачка, а нижняя часть (влагальце) архегония окружает основание ножки воротничком.
Колпачок — плёнчатое образование, покрывающее молодую коробочку сверху целиком или частично, защищая её от внешних воздействий. Колпачок обычно плотный, сверху по большей части многослойный. Суженная буроватая верхушка колпачка часто соответствует шейке архегония. Обертка зародыша, называемая эпигон, первоначально цельная, формируется за счёт архегония, его ножки, и, отчасти, стебля.
У большинства листостебельных мхов колпачок хорошо развит, не считая некоторых клейстокарпных эфемеров из бриевых. По форме колпачки могут быть очень разнообразны. Колпачок обычно опадает ещё до полного развития коробочки, реже он долго остается на коробочке и опадает вместе с крышечкой.
_8522.JPG)
У большинства бриевых ножка развита хорошо, но длина её широко варьирует даже у представителей одного семейства. У некоторых родов бриевых ножка полностью отсутствует или присутствует, но не достигает полного развития. Обычно так обстоит дело у эфемеров с коробочками, закрытыми неотделяющейся крышечкой, и погруженными в перихециальные листья. У многих видов ткани ножки спорогония те же, что и в стебле гаметофита. Снаружи ножки располагаются толстостенные клетки механической ткани, затем паренхима коры, в центре проходит тяж из проводящих прозенхимных клеток.
Ножка большинства листостебельных мхов очень чувствительна к колебаниям влажности воздуха. Подсыхая, ножка спирально закручивается и толчками вращает коробочку, активно рассеивая споры. Наружные стереидные клетки при этом тоже расположены по спирали. Причина гигроскопического движения ножки — способность наружных стенок стереид быстро впитывать влагу, разбухать и раскручивать ножку, а при высыхании, наоборот, быстро терять воду и скручивать ножку.
Стопа, то есть нижняя часть ножки, уходит глубоко в ткань гаметофита и прикрепляет к нему спорогон. Основная функция стопы — получение из гаметофита необходимых для развития плода веществ. У одних мхов стопа состоит из однородных клеток, у других поверхностные клетки вытягиваются в сосочки наподобие корневых волосков. Иногда они даже порождают разветвлённые многоклеточные ризоиды. Эти ризоиды активно поглощают воду и питательные вещества. Центральный пучок ножки может продолжаться и дальше в стопу, а нередко даже проникает в центральный пучок стебля.
Влагальце передаёт питательные вещества от гаметофита к растущему спорофиту через стопу. Гаметофит и спорофит не образуют прочной связи, спорофит легко отделяется от гаметофита.
_7673.JPG)
Коробочка состоит из крышечки, колечка, урны, шейки и апофизы, если последняя вообще есть. У некоторых видов коробочки имеют неотделяющуюся крышечку. Такие мхи называются клейстокарпными (закрытоплодными). Мхи с отделяющейся крышечкой называются стегокарпными (крышкоплодными). Крышечка прикрывает собой устье коробочки и отпадает после созревания спор. В сбрасывании крышечки участвуют колечко и зубцы перистома. Ко времени созревания коробочки ткани колечка значительно натягиваются, помогая сбросить крышечку.
_5491.JPG)
Колечко располагается между верхним краем урны и крышечкой. Оно состоит из лежащих друг над другом в один или несколько рядов уплощённых бесцветных, сильно гигроскопичных клеток. При набухании этих клеток колечко расширяется и отделяется и от урны, и от крышечки. Колечко представляет собой полоску в виде пояса, клетки его крупные.
Урной называют часть коробочки, внутри которой развиваются споры. В верхней части урны находится широкое или суженное отверстие — устье, закрытое крышечкой. По краю устья часто формируются выросты в виде зубцов разнообразной формы, которые называются перистомом или околоустьем.
Стенки коробочки состоят из двух или нескольких слоев. Наружный слой — экзотеций, гомологичный эпидерме высших растений, кутинизирован. Клетки экзотеция бедны плазмой, их наружные оболочки обычно утолщены и плотно прилегают друг к другу. Внутренние слои стенки коробочки состоят из крупных паренхиматических клеток с тонкими бесцветными клеточными оболочками, они образуют водоносную ткань.
Клетки стенок молодого спорогона богаты хлоропластами, по мере созревания число хлоропластов уменьшается. Экзотеций перестает выполнять ассимиляционную функцию, оставляя за собой только механическую. В экзотеции нижней половины урны и шейки находятся устьица. Поверхностные устьица располагаются на уровне экзотеция, погруженные устьица — расположены ниже его уровня. Устьица обычной формы из двух симметричных почковидных замыкающих клеток с щелью. У бриевых из родов архидиум, схистостега, тетрафис устьица отсутствуют совсем.
_9880.JPG)
Под экзотецием лежит однослойная, двуслойная или четырёхслойная водоносная ткань. Клетки водоносной ткани крупнее клеток экзотеция, они тонкостенные и заполнены водой. Водоносная ткань снабжает водой лежащую ещё глубже ассимиляционную ткань. Благодаря тонким стенкам и прозрачности клеток водоносная ткань пропускает световые лучи к хлоропластам.
У большинства мхов между стенками коробочки и споровым мешком находится воздушная полость, пронизанная нитями из хлорофиллоносных клеток. Воздушная полость и ассимиляционная ткань отсутствуют у родов, где споровый мешок непосредственно примыкает к стенке коробочки, а также у водных мхов.
В центре урны располагается колонка. Она состоит из прозенхимных клеток, проводящих к развивающимся спорам воду и питательные вещества. Колонка обычно проходит в коробочке от шейки до крышечки. У немногих видов она вытягивается, выступает наружу из урны и поднимает оставшуюся прикреплённой к ней крышечку. После отпадения крышечки у многих мхов колонка сморщивается и остается на дне урны.
Перистом состоит из зубцов, расположенных а один или два ряда, из ресничек или нитей разнообразной формы. У некоторых мхов перистом отсутствует, у других он — недоразвитый, рудиментарный.

Зубцы однорядного перистома и экзостома (наружного ряда) двойного перистома способны к гигроскопическому движению. Они состоят из двух пластин, имеющих разную анатомическую структуру, и, соответственно, различную степень гигроскопичности. Движения зубцов активно помогают высеивать споры из урны и распространять их как можно шире. Также этими движениями зубцы закрывают устье урны при высокой влажности воздуха, не давая спорам намокнуть и прорасти внутри коробочки.
Нижняя часть коробочки, не содержащая в себе спор, шейка, постепенно или внезапно переходит в ножку. Нередко ткань на шейке односторонне разрастается, формируя зобик. Зобики характерны для некоторых мхов из дикрановых.
Апофиза — это вздутие нижней части коробочки, принимающее разнообразные формы. Яркая окраска апофизы и выделяемая ею через устьица жидкость с неприятным запахом привлекает к себе мух, которые становятся распространителями спор.
В разделе не хватает ссылок на источники (см. рекомендации по поиску). |
Размножение
правитьСпоры
правитьСпоры листостебельных мхов обычно представляют собой одноклеточные образования, содержащие хлоропласты и капли масла. Размеры их сильно колеблются, но чаще всего имеют диаметр 10-12 мкм. У некоторых родов, преимущественно тропических, споры многоклеточные, начинающие прорастать, ещё находясь в спорангии.
Обычно споры листостебельных мхов шаровидные, реже овальные, иногда слабо округло-угловатые, редко почковидные.
Споры мхов очень жизнеспособны, могут переносить как засухи, так и холода. Сухие споры после замораживания их при температуре ниже — 200 °C в течение нескольких часов нормально прорастали на питательных средах. Хорошо выносят кратковременное нагревание до +100 °C.
При оптимальных условиях освещения, температуры, влажности и характерной для данного вида реакции субстрата споры прорастают обычно через несколько дней или недель. Из прорастающих спор развивается первичная протонема, на которой закладываются почки. Из почек вырастают молодые растения.
При набухании споры экзина (внешняя прочная оболочка) лопается, а интина вытягивается в виде сосочка вместе с содержимым споры. Делясь, оно даёт начало либо однорядной нити, либо однослойной (редко многослойной) пластинке, несущей ризоиды. Эта стадия развития гаметофита называется протонемой. Первая клетка гаметофита прикрыта только интиной (тонкой плёнкой из целлюлозы и пектина) и беззащитна перед влиянием внешней среды. При малейших неблагоприятных условиях она обречена на гибель. Протонема состоит из зелёной хлоронемы (фотосинтезирующей) и подземной части — бесцветной ризонемы. Величина, форма и продолжительность жизни протонемы у листостебельных мхов значительно варьируют. Ветвящаяся протонема некоторых видов мхов покрывает площадь до 1 м2 и существует месяцами. У видов с однолетними небольшими гаметофитами протонема может жить несколько лет. Обычно же размеры протонемы достигают нескольких сантиметров, а продолжительность жизни составляет несколько дней или недель.

Вегетативное размножение
правитьВторичную протонему в виде нити, на которой образуются почки, а затем молодые растения, или специальные приспособления для вегетативного размножения (выводковые тельца, листья, почки и т. д.) могут сформировать почти любые части гаметофита листостебельных мхов.
По мере разрастания моховой дернины от неё обособляются молодые побеги в тот момент, когда нижняя часть материнского растения отмирает. Это самый распространённый способ вегетативного размножения мхов.
Возможно размножение частями тела — обломками стеблей, почками и ветвями, выводковыми ветвями, выводковыми почками (укороченными выводковыми ветвями с нередуцированными или с редуцированными листьями), обломками листьев, выводковыми листьями.
Выводковые листья отделяются целиком и обычно отличаются от других листьев. Иногда выводковые листья собраны в головки на конце ножки.
Выводковые нити, как и почки, образуются обычно в пазухах. Представляют собой нитевидные образования, иногда разветвлённые, часто окрашенные, гладкие или паппилозные.
Кроме того, на любых частях растения могут возникать выводковые тела — многоклеточные образования для вегетативного размножения. Они образуются на вторичной протонеме, на стеблевом ризоидном войлоке, на стебле в пазухах листьев, на различных частях жилки листа, на клетках листовой пластинки. К моменту отделения выводкового тела в месте его соединения с материнским растением образуется разделительная клетка с очень нежной наружной стенкой, которая легко разрушается. По наиболее частым местам образования различают стеблевые, листовые и пазушные выводковые тела. Выводковые тела могут иметь различную форму и окраску, быть как одиночными, так и скученными.
Выводковые органы распространяются потоками воздуха, водой, разносятся животными.
Классификация
правитьВ прошлом в группу Bryopsida были включены все мхи. Текущий состав группы является более ограниченным.
В классификации, предложенной Goffinet B. и W. R. Buck в 2006 году, класс включает 6 подклассов[1][2]:
- подкласс Бриевые (Bryidae) (71 семейство)
- подкласс Буксбаумиевые (Buxbaumiidae) (единственный род Буксбаумия (Buxbaumia))
- подкласс Dicranidae (24 семейства)
- подкласс Diphysciidae (единственный род Дифисциум (Diphyscium))
- подкласс Funariidae (5 семейств)
- подкласс Timmiidae (единственный род Timmia)
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Текущая филогения класса Bryopsida. | |||||||||||||||||||||||||||||||||||||||||||||||||||
Более детальная филогения до уровня порядков по работе Новикова и Барабаш-Красны 2015 года. [3]
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Согласно базе данных Catalogue of Life[4] на май 2023 года класс включает следующие подклассы:
- Bryidae Engl., 1892 — Бриевые
- Buxbaumiidae Doweld., 2001 — Буксбаумиевые
- Dicranidae Doweld., 2001
- Diphysciidae Ochyra, 2003
- Funariidae Ochyra, 2003
- Gigaspermidae M. Stech et W. Frey, 2008
- Timmiidae Ochyra, 2003
Литература
править- Жизнь растений. Том 4. Мхи. Планктоны. Хвощи. Папоротники. Голосеменные растения // Под ред. И. В. Грушивицкого, С. Г. Жилина. — М.: Просвещение, 1978. — 447 с.
- Слука З. А. Зелёные мхи. — М.: Издательство Московского университета, 1980. — 134 с.
Примечания
править- ↑ Goffinet B., Buck W. R. & Shaw A. J. (2008). «Morphology and Classification of the Bryophyta» in Goffinet B. & Shaw J. (eds.) Bryophyte Biology, 2nd ed. (New York: Cambridge University Press). pp. 55—138. ISBN 978-0-521-87225-6.
- ↑ Goffinet, Bernard; William R. Buck. Systematics of the Bryophyta (Mosses): From molecules to a revised classification (англ.) // Monographs in Systematic Botany : journal. — Missouri Botanical Garden Press, 2004. — Vol. 98. — P. 205—239. — ISBN 1-930723-38-5.
- ↑ Novíkov & Barabaš-Krasni. Modern plant systematics (неопр.). — Liga-Pres, 2015. — С. 685. — ISBN 978-966-397-276-3. — doi:10.13140/RG.2.1.4745.6164.
- ↑ Catalogue of Life. Дата обращения: 4 мая 2023. Архивировано 5 октября 2018 года.