Ящеротазовые
Ящерота́зовые[2], или ящерота́зовые диноза́вры, или заври́схии[3] (лат. Saurischia), — одна из двух традиционно выделяемых основных клад динозавров (наряду с птицетазовыми)[4][5][6][7], исторически рассматриваемая в ранге отряда[8][9]. У ящеротазовых, в отличие от птицетазовых, лобковые кости первично наклонены вперёд (у некоторых представителей — герреразаврид[10], теризинозавроид[11], дромеозаврид[12] и птиц[13] — лобковые кости отклоняются назад). Вопреки названию, ящеротазовые, а не птицетазовые, дали начало птицам, которые, по преобладающим среди палеонтологов позвоночных[англ.] кладистическим представлениям, являются единственной пережившей мел-палеогеновое вымирание группой ящеротазовых[4][5][6][7].
| Ящеротазовые | |||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 1-й ряд (ранние ящеротазовые): апатозавр A. louisae и платеозавр P. engelhardti | |||||||||||||||||||||||||||||||||||||||||
| Научная классификация | |||||||||||||||||||||||||||||||||||||||||
|
Домен: Царство: Подцарство: Без ранга: Без ранга: Тип: Подтип: Инфратип: Надкласс: Клада: Клада: Клада: Клада: Клада: Клада: Клада: Ящеротазовые |
|||||||||||||||||||||||||||||||||||||||||
| Международное научное название | |||||||||||||||||||||||||||||||||||||||||
| Saurischia Seeley, 1888 | |||||||||||||||||||||||||||||||||||||||||
| Синонимы | |||||||||||||||||||||||||||||||||||||||||
|
Источник: [1]
|
|||||||||||||||||||||||||||||||||||||||||
| Классификация | |||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||
| Геохронология появился 235 млн лет
◄ Наше время
◄ Мел-палеогеновое вымирание ◄ Триасовое вымирание
◄ Массовоепермское вымирание ◄ Девонское вымирание
◄ Ордовикско-силурийское вымирание
◄ Кембрийский взрыв
|
|||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||
Ящеротазовые обычно считаются монофилетической («естественной») группой, включающей обширные клады теропод (Theropoda) и завроподоморф (Sauropodomorpha) в составе группы Eusaurischia («настоящие ящеротазовые»)[4][6], а также боковую базальную ветвь Herrerasauria (герреразавриды и родственники)[6][14][15], которую иногда помещают в состав теропод[16]. Согласно альтернативной точке зрения, тероподы более близки к птицетазовым, чем к завроподоморфам, и, соответственно, ящеротазовые в традиционном понимании являются парафилетической («искусственной») группой, которую следует либо расформировать, либо переопределить (последнее радикально изменило бы её состав)[17][18][19].
Описание
править
1-й ряд: дейнохейрус и тарбозавр, белая сова;
2-й ряд: чжэньюаньлун[англ.][20], "Coelophysis" kayentakatae;
3-й ряд: синозавроптерикс, спинозавр.

.jpg)
Древнейшими представителями ящеротазовых были такие динозавры, как герреразавриды герреразавр[21] и ставрикозавр[22], завроподоморф эораптор[23] и теропод эодромей[24], жившие примерно 235—228 млн лет назад (карнийский век, верхний триас)[25]. Судя по ископаемым остаткам этих животных и их близких родственников, ранние представители клады являлись небольшими неспециализированными плотоядными, передвигавшимися на двух конечностях[21][22][23][24].
Ранние тероподы имели довольно консервативную анатомию, общие черты которой были характерны для предков динозавров. Облигатно бипедальная походка предков динозавров была сохранена всеми тероподами; большинство нептичьих теропод обладало уплощёнными с боков зубами с режущими пильчатыми краями, называемыми зифодонтными, что говорит об их плотоядности[26]. В процессе эволюции некоторые тероподы группы целурозавров, такие как теризинозавры, орнитомимозавры, овирапторозавры, альваресзавриды и авиалы (птицы), адаптировались к всеядности или растительноядности, что позволило им освоить новые экологические ниши[27]. Тероподы, как ранние, так и продвинутые, сильно варьировали в размерах. По оценкам, длина спинозавра, одного из крупнейших теропод (возможно, крупнейшего), могла достигать до 16 м. Длина современной колибри-пчёлки составляет примерно 5 см (энанциорнисовая птица Cratoavis была ненамного больше — 6,6 см в длину); некоторые нептичьи тероподы, такие как Epidexipteryx, достигали около 25 см в длину[28].
Первые завроподоморфы постепенно перешли с хищничества на всеядность, а позже и на растительноядность[29], при переходе к завроподам сменив бипедальную походку на квадропедальную[30][31]. Последние завроподоморфы, не относящиеся к завроподам (ранее выделялись в группу прозавропод), вымерли в ранней юре[7]. Завроподы были квадропедальными растительноядными со столбообразными конечностями и длинными хвостами и шеями. Сочетание унаследованных от предков признаков и эволюционных новшеств позволило представителям разных групп завропод поздней юры и мелового периода независимо вырасти до гигантских размеров (хотя известны и карликовые формы[32][33])[34]. Тогда как первые завроподоморфы достигали около 1,5 м в длину, длина крупнейших завропод могла достигать 35—45 м, что делает их самыми длинными когда-либо существовавшими позвоночными[35][36].
Последние ящеротазовые, не относящиеся к веерохвостым птицам, вымерли вместе с птицетазовыми и многими другими группами в ходе мел-палеогенового вымирания 66 млн лет назад. Предполагается, что выживание веерохвостых птиц обеспечила их адаптация к поеданию семян[англ.], оказавшаяся жизненно важной в условиях вымирания[37][38].
-
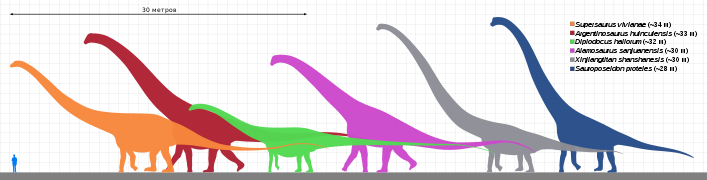 Некоторые гигантские завроподы
Некоторые гигантские завроподы -
 Некоторые гигантские тероподы
Некоторые гигантские тероподы -
 Ранний завроподоморф Buriolestes
Ранний завроподоморф Buriolestes -
 Одни из самых маленьких известных нептичьих теропод
Одни из самых маленьких известных нептичьих теропод -
 Колибри-пчёлка является самым маленьким известным науке ящеротазовым
Колибри-пчёлка является самым маленьким известным науке ящеротазовым




.svg)
Систематика
править

В 1888 году была опубликована работа британского палеонтолога Гарри Говира Сили, где он изложил свои выводы о том, что динозавры не являются единой эволюционной группой (монофилия), как это было принято тогда считать, а состоят из двух независимо возникших ветвей (полифилия). Сили указал в качестве ключевого различия между этими ветвями строение таза: у ящеротазовых лобковые кости первично наклонены вперёд, как у современных ящериц и крокодилов, а у птицетазовых — назад, как у птиц[8]. Представление о полифилии динозавров имело широкое распространение в научных кругах вплоть до начавшегося в 1960-х годах «Ренессанса динозавров», в ходе которого было опровергнуто[39][40]. Большинство современных авторов согласно с тем, что птицетазовые и ящеротазовые являются двумя основными подгруппами монофилетической группы динозавров[4][5][6][7].
Ящеротазовые с видоизменённым строением таза


Названия «ящеротазовые» и «птицетазовые» носят весьма условный характер, поскольку Сили дал их, не предполагая происхождение птиц от ящеротазовых и вообще их связь с динозаврами. В настоящее время общепринято, что птицы произошли от ящеротазовых с типичным для своей группы тазом, но конвергентно с птицетазовыми приобрели таз с лобковой костью, наклоненной назад. Таким образом, в соответствии с принципами кладистики, птицы являются единственной ныне живущей группой ящеротазовых динозавров, тогда как птицетазовые полностью вымерли, не оставив потомков[41].
Согласно филогенетическому определению, рекомендуемому «Филокодексом» (Gauthier et al., 2020), название Saurischia соответствует наибольшей кладе, включающей Allosaurus fragilis Marsh, 1877 (Theropoda) и Camarasaurus supremus Cope, 1877 (Sauropodomorpha), но не Stegosaurus stenops Marsh, 1887 (Ornithischia)[5][42]. Существует также ряд определений, где в качестве таксона-спецификатора ящеротазовых указывается только теропод[6][43], либо только завроподоморф[17]; проблемой таких определений является то, что если Saurischia в традиционном понимании не являются монофилетической группой, то этот таксон окажется синонимом Sauropodomorpha или Theropoda, либо очень близким к ним по составу (тогда как определение «Филокодекса» подразумевает, что таксон будет просто расформирован[5]).
По составу с Saurischia схож таксон Eusaurischia (буквально — «настоящие ящеротазовые»), введённый К. Падианом[англ.], Дж. Р. Хатчинсоном и Т. Р. Хольцем в 1999 году для обозначения наиболее инклюзивной клады, включающей завроподоморф и теропод, то есть всех ящеротазовых, за исключением некоторых базальных таксонов (возможно, в число этих таксонов входят Herrerasauria)[44]. М. К. Лангер (2004) определил Eusaurischia как наиболее инклюзивную кладу, включающую цетиозавра (Cetiosaurus) и веерохвостых птиц (Neornithes)[6].
Филогения и альтернативные классификации
править| Филогенетические взаимоотношения между основными группами динозавров[45] | |||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
До того как Сили разделил динозавров на птицетазовых и ящеротазовых, существовали и другие системы классификации. В 1870 году Томас Гексли выделил новый таксон орнитосцелид (Ornithoscelida) для объединения компсогната (в группе Compsognatha) и динозавров (Dinosauria)[46], в состав которых на тот момент включали таксоны, ныне рассматриваемые как нептичьи тероподы и птицетазовые. Хотя с тех пор «орнитосцелидная» гипотеза уступила место классификации Сили, она была возрождена в 2017 году М. Бароном и соавторами, филогенетический анализ которых восстановил теропод в качестве сестринского таксона птицетазовых, а не завроподоморф[17]. Результаты анализа не были приняты всеми авторами, и в настоящее время филогенетические взаимоотношения между основными группами динозавров всё ещё обсуждаются[5][15][18][47][48].
М. Барон и соавторы (2017) определили Saurischia как наиболее инклюзивную кладу, включающую диплодока Карнеги (Diplodocus carnegii), но не трицератопса грубого (Triceratops horridus). В соответствии с их филогенетическим анализом, под это определение попадают завроподоморфы и герреразавриды, но не тероподы, что подразумевает радикальный пересмотр состава клады ящеротазовых[17]. По мнению Томаса Хольца, если завроподоморфы и тероподы не являются сестринскими таксонами, то название Saurischia следует вывести из употребления[49].
В 1986 году Р. Бэккер предположил, что завроподоморфы могут быть более близки к птицетазовым, чем к тероподам, и объединил их в кладу фитодинозавров[англ.][45] (Phytodinosauria)[50]. Единственным анализом, восстановившим такую топологию, является анализ Л. Пэрри и соавторов (2017), причём данный результат был получен только при определённых критериях оптимальности[51].
Ещё до того как Сили выделил ящеротазовых, в 1860 году, Ричард Оуэн, известный как автор таксона Dinosauria (динозавры), выделил группу Opisthocoelia для объединения Cetiosaurus (завроподоморфы) и Streptospondylus (теропод), которых он рассматривал как крокодилов[52]. В 1914 году Фридрих фон Хюне выделил таксон Pachypodosauria для объединения завроподоморф и карнозавров (в его понимании это были любые крупные тероподы), но не целурозавров (в его понимании — любые мелкие тероподы)[53]. Большинство таксонов, включённых в состав Opisthocoelia и Pachypodosauria, теперь рассматривается как ящеротазовые клады Eusaurischia[1].
| Müller & Garcia, 2020[54] | Novas et al., 2021[55] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| «Орнитосцелидная» гипотеза. Таксоны, традиционно относимые к ящеротазовым, выделены зелёным фоном. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Baron et al., 2017[17] | Cau, 2018[19] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Примечания
править- ↑ 1 2 Mortimer M. Saurischia (англ.). The Theropod Database. Дата обращения: 17 мая 2021. Архивировано 10 октября 2021 года.
- ↑ Еськов К. Ю. Словарь названий живых и ископаемых организмов // Удивительная палеонтология: история Земли и жизни на ней. — М. : ЭНАС, 2008. — 312 с. — ISBN 978-5-91921-129-7.
- ↑ Татаринов, 2009, с. 68.
- ↑ 1 2 3 4 Saurischia (англ.). Encyclopædia Britannica. Дата обращения: 17 мая 2021.
- ↑ 1 2 3 4 5 6 de Queiroz, Cantino & Gauthier, 2020, Saurischia H. G. Seeley 1888 [J. A. Gauthier, M. C. Langer, F. E. Novas, J. Bittencourt & M. D. Ezcurra], converted clade name, pp. 1209—1217.
- ↑ 1 2 3 4 5 6 7 Weishampel, Dodson & Osmolska, 2004, "Saurischia" by Holtz T. R. Jr. and Osmolska H., pp. 21—24.
- ↑ 1 2 3 4 Benton, 2015, chpt. 8.3: "The Diversity Of Saurischian Dinosaurs", pp. 207—221.
- ↑ 1 2 Seeley H. G. I. On the classification of the fossil animals commonly named Dinosauria (англ.) // Proceedings of the Royal Society of London : journal. — 1888. — Vol. 43, iss. 258—265. — P. 165—171. — doi:10.1098/rspl.1887.0117. Архивировано 18 мая 2021 года.
- ↑ Benton, 2015, p. 440.
- ↑ Paul, 1988, pp. 248—250.
- ↑ Weishampel, Dodson & Osmolska, 2004, chpt. 7: "Therizinosauroidea" by J. M. Clark, T. Maryanska[англ.], and R. Barsbold, pp. 151—164.
- ↑ Weishampel, Dodson & Osmolska, 2004, chpt. 10: "Dromaeosauridae" by P. J. Makovicky and M. A. Norell[англ.], pp. 196—210.
- ↑ Weishampel, Dodson & Osmolska, 2004, chpt. 11: "Basal Avialae" by K. Padian[англ.], pp. 210—231.
- ↑ Novas F. E.[англ.], Agnolin F. L., Ezcurra M. D.[англ.], Müller R. T., Martinelli A., Langer M. Review of the fossil record of early dinosaurs from South America, and its phylogenetic implications (англ.) // Journal of South American Earth Sciences[англ.]. — 2021. — P. 103341. — ISSN 0895-9811. — doi:10.1016/j.jsames.2021.103341.
- ↑ 1 2 Langer M. C., Ezcurra M. D.[англ.], Rauhut O. W. M., Benton M. J.[англ.], Knoll F. Untangling the dinosaur family tree (англ.) // Nature : journal. — 2017. — Vol. 551, iss. 7678. — P. E1—E3. — ISSN 1476-4687. — doi:10.1038/nature24011. Архивировано 10 октября 2021 года.
- ↑ Nesbitt S. J.[англ.]. The Early Evolution of Archosaurs: Relationships and the Origin of Major Clades (англ.) // Bulletin of the American Museum of Natural History[англ.] : journal. — 2011. — Vol. 2011, iss. 352. — P. 1—292. — ISSN 1937-3546 0003-0090, 1937-3546. — doi:10.1206/352.1. Архивировано 1 июля 2019 года.
- ↑ 1 2 3 4 5 Baron M. G., Norman D. B., Barrett P. M. A new hypothesis of dinosaur relationships and early dinosaur evolution (англ.) // Nature : journal. — 2017. — Vol. 543, iss. 7646. — P. 501—506. — ISSN 1476-4687. — doi:10.1038/nature21700.
- ↑ 1 2 Baron M. G., Norman D. B., Barrett P. M. Baron et al. reply (англ.) // Nature : journal. — 2017. — Vol. 551, iss. 7678. — P. E4—E5. — ISSN 1476-4687. — doi:10.1038/nature24012.
- ↑ 1 2 Cau A. The assembly of the avian body plan: a 160-million-year long process (англ.) // Bollettino della Societa Paleontologica Italiana : journal. — 2018. — Vol. 57, iss. 1. — P. 1–25. — ISSN 0375-7633. — doi:10.4435/BSPI.2018.01. Архивировано 21 декабря 2018 года.
- ↑ Брусатти, 2019, с. 14.
- ↑ 1 2 Novas F. E.[англ.]. New information on the systematics and postcranial skeleton of Herrerasaurus ischigualastensis (Theropoda: Herrerasauridae) from the Ischigualasto Formation (Upper Triassic) of Argentina (англ.) // Journal of Vertebrate Paleontology : journal. — 1994. — Vol. 13, iss. 4. — P. 400—423. — ISSN 0272-4634. — doi:10.1080/02724634.1994.10011523. Архивировано 18 мая 2021 года.
- ↑ 1 2 Bittencourt J. S., Kellner A. W. A. The anatomy and phylogenetic position of the Triassic dinosaur Staurikosaurus pricei Colbert, 1970 (англ.) // Zootaxa : journal. — 2009. — Vol. 2079, iss. 1. — P. 1—56. — ISSN 1175-5334. — doi:10.11646/zootaxa.2079.1.1. Архивировано 18 мая 2021 года.
- ↑ 1 2 Sereno P. C., Martinez R. N., Alcober O. A. Osteology of Eoraptor lunensis (Dinosauria, Sauropodomorpha) (англ.) // Journal of Vertebrate Paleontology : journal. — 2012. — Vol. 32, iss. sup1. — P. 83—179. — ISSN 0272-4634. — doi:10.1080/02724634.2013.820113. Архивировано 11 мая 2021 года.
- ↑ 1 2 Martinez R. N., Sereno P. C., Alcober O. A., Colombi C. E., Renne P. R. A Basal Dinosaur from the Dawn of the Dinosaur Era in Southwestern Pangaea (англ.) // Science : journal. — 2011. — Vol. 331, iss. 6014. — P. 206—210. — ISSN 1095-9203 0036-8075, 1095-9203. — doi:10.1126/science.1198467. Архивировано 18 мая 2021 года.
- ↑ Holtz T. R. Jr. Holtz's Genus LIst Архивная копия от 2 марта 2012 на Wayback Machine (англ.). — 2012. — P. 2; 24.
- ↑ Weishampel, Dodson & Osmolska, 2004, "Saurischia" by Holtz T. R. Jr. and Osmolska H., pp. 21—24.
- ↑ Zanno L. E.[англ.], Makovicky P. J. Herbivorous ecomorphology and specialization patterns in theropod dinosaur evolution (англ.) // Proceedings of the National Academy of Sciences : journal. — 2011. — Vol. 108, iss. 1. — P. 232—237. — ISSN 1091-6490 0027-8424, 1091-6490. — doi:10.1073/pnas.1011924108.
- ↑ Molina-Pérez & Larramendi, 2019, pp. 72—75.
- ↑ Martinez R. N., Alcober O. A. A Basal Sauropodomorph (Dinosauria: Saurischia) from the Ischigualasto Formation (Triassic, Carnian) and the Early Evolution of Sauropodomorpha (англ.) // PLoS ONE : journal. — 2009. — Vol. 4, iss. 2. — P. 1—12. — ISSN 1932-6203. — doi:10.1371/journal.pone.0004397. Архивировано 24 сентября 2015 года.
- ↑ Yates A. M., Bonnan M. F., Neveling J., Chinsamy A., Blackbeard M. G. A new transitional sauropodomorph dinosaur from the Early Jurassic of South Africa and the evolution of sauropod feeding and quadrupedalism (англ.) // Proceedings of the Royal Society B: Biological Sciences : journal. — 2010. — Vol. 277, iss. 1682. — P. 787—794. — ISSN 1471-2954. — doi:10.1098/rspb.2009.1440. — PMID 19906674. Архивировано 28 мая 2022 года.
- ↑ Otero A., Cuff A. R., Allen V., Sumner-Rooney L., Pol D. Ontogenetic changes in the body plan of the sauropodomorph dinosaur Mussaurus patagonicus reveal shifts of locomotor stance during growth (англ.) // Scientific Reports[англ.] : journal. — 2019. — Vol. 9, iss. 1. — P. 7614. — ISSN 2045-2322. — doi:10.1038/s41598-019-44037-1. Архивировано 28 сентября 2021 года.
- ↑ Sander P. M., Mateus O. V., Laven T., Knotschke N. Bone histology indicates insular dwarfism in a new Late Jurassic sauropod dinosaur (англ.) // Nature : journal. — 2006. — Vol. 441, iss. 7094. — P. 739—741. — ISSN 1476-4687. — doi:10.1038/nature04633. — . — PMID 16760975. Архивировано 18 мая 2021 года.
- ↑ Stein K., Csiki Z., Rogers K. C.[англ.], Weishampel D. B., Redelstorff R. Small body size and extreme cortical bone remodeling indicate phyletic dwarfism in Magyarosaurus dacus (Sauropoda: Titanosauria) (англ.) // PNAS : journal. — 2010. — Vol. 107, iss. 20. — P. 9258–9263. — ISSN 0027-8424. — doi:10.1073/pnas.1000781107. — PMID 20435913. Архивировано 27 мая 2020 года.
- ↑ Sander P. M., Christian A., Clauss M., Fechner R., Gee C. T. Biology of the sauropod dinosaurs: the evolution of gigantism (англ.) // Biological Reviews of the Cambridge Philosophical Society : journal. — 2011. — Vol. 86, iss. 1. — P. 117—155. — ISSN 1464-7931. — doi:10.1111/j.1469-185X.2010.00137.x. Архивировано 20 мая 2013 года.
- ↑ Molina-Pérez & Larramendi, 2020, p. 54—59.
- ↑ Paul G. S.[англ.]. Determining the largest known land animal: A critical comparison of differing methods for restoring the volume and mass of extinct animals (англ.) // Annals of the Carnegie Museum[англ.] : journal. — 2019. — Vol. 85, no. 4. — P. 335—358. — ISSN 1943-6300. — doi:10.2992/007.085.0403. Архивировано 15 ноября 2021 года.
- ↑ Field D. J., Bercovici A., Berv J. S., Dunn R., Fastovsky D. E., Lyson T. R., Vajda V., Gauthier J. A. Early Evolution of Modern Birds Structured by Global Forest Collapse at the End-Cretaceous Mass Extinction (англ.) // Current Biology : journal. — 2018. — Vol. 28, iss. 11. — P. 1825—1831. — ISSN 0960-9822. — doi:10.1016/j.cub.2018.04.062. Архивировано 16 сентября 2023 года.
- ↑ Larson D. W., Brown C. M., Evans D. C.[англ.]. Dental Disparity and Ecological Stability in Bird-like Dinosaurs prior to the End-Cretaceous Mass Extinction (англ.) // Current Biology : journal. — 2016. — Vol. 26, iss. 10. — P. 1325—1333. — ISSN 0960-9822. — doi:10.1016/j.cub.2016.03.039.
- ↑ Novas F. E.[англ.]. Dinosaur Monophyly (англ.) // Journal of Vertebrate Paleontology : journal. — 1996. — Vol. 16, iss. 4. — P. 723—741. — ISSN 0272-4634.
- ↑ Bakker R. T., P. M. Galton[англ.]. Dinosaur Monophyly and a New Class of Vertebrates (англ.) // Nature : journal. — 1974. — Vol. 248, iss. 5444. — P. 168—172. — ISSN 1476-4687. — doi:10.1038/248168a0. Архивировано 18 мая 2021 года.
- ↑ Brusatte, 2012, pp. 20—21.
- ↑ Saurischia (англ.). RegNum. Дата обращения: 17 мая 2021. Архивировано 18 мая 2021 года.
- ↑ Brett-Surman, Holtz & Farlow, 2012, chpt. 11: "The Taxonomy and Systematics of the Dinosaurs" by Holtz T. R. Jr. and M. K. Brett-Surman, p. 236—251.
- ↑ Padian K.[англ.], Hutchinson J. R., Holtz T. R. Jr. Phylogenetic definitions and nomenclature of the major taxonomic categories of the carnivorous Dinosauria (Theropoda) (англ.) // Journal of Vertebrate Paleontology : journal. — 1999. — Vol. 19, iss. 1. — P. 69—80. — ISSN 0272-4634. — doi:10.1080/02724634.1999.10011123. Архивировано 18 мая 2021 года.
- ↑ 1 2 Нэйш и Барретт, 2019, с. 41.
- ↑ Huxley T. H. On the Classification of the Dinosauria, with observations on the Dinosauria of the Trias (англ.) // Quarterly Journal of the Geological Society : journal. — 1870. — Vol. 26, iss. 1—2. — P. 32—51. — ISSN 0370-291X. — doi:10.1144/GSL.JGS.1870.026.01-02.09. Архивировано 18 мая 2021 года.
- ↑ Kirmse J. P. D. Morphological support of Ornithoscelida Huxley, 1870 (англ.) // Master's Dissertation, Faculdade de Filosofia, Ciências e Letras de Ribeirão Preto, University of São Paulo, Ribeirão Preto. — 2021. — P. 1—300. — doi:10.11606/D.59.2021.tde-24082021-141444. Архивировано 12 сентября 2021 года.
- ↑ "A discussion, not a war: two opposing experts talk dinosaur family trees". The Guardian (англ.). 2018. Архивировано 9 ноября 2020. Дата обращения: 15 мая 2021.
- ↑ 1 2 Holtz T. R. Jr. Share names for dinosaur divisions (англ.) // Nature : journal. — 2017. — Vol. 545, iss. 7652. — P. 30. — ISSN 1476-4687. — doi:10.1038/545030d. Архивировано 19 октября 2021 года.
- ↑ Bakker, 1986, p. 459.
- ↑ Parry L. A., Baron M. G., Vinther J. Multiple optimality criteria support Ornithoscelida (англ.) // Royal Society Open Science[англ.] : journal. — 2017. — Vol. 4, iss. 10. — ISSN 2054-5703. — doi:10.1098/rsos.170833. Архивировано 12 ноября 2020 года.
- ↑ Owen, 1860, p. 272—273.
- ↑ F. v. Huene. Beitrage zur geschichte der Archosaurier (нем.) // Geologie und Palaontologie Abhandlungen. — 1914. — Bd. 13, Nr. 7. — S. 1—56.
- ↑ Müller R. T., Garcia M. S. A paraphyletic ‘Silesauridae' as an alternative hypothesis for the initial radiation of ornithischian dinosaurs (англ.) // Biology Letters : journal. — 2020. — Vol. 16, iss. 8. — P. 20200417. — ISSN 1744-957X. — doi:10.1098/rsbl.2020.0417. — PMID 32842895. Архивировано 29 августа 2022 года.
- ↑ Novas F. E.[англ.], Agnolin F. L., Ezcurra M. D.[англ.], Müller R. T., Martinelli A., Langer M. Review of the fossil record of early dinosaurs from South America, and its phylogenetic implications (англ.) // Journal of South American Earth Sciences[англ.]. — 2021. — P. 103341. — ISSN 0895-9811. — doi:10.1016/j.jsames.2021.103341.
Литература
правитьНа русском языке
править- Брусатти С. Время динозавров. Новая история древних ящеров = The Rise and Fall of the Dinosaurs: A New History of a Lost World / науч. ред. А. О. Аверьянов. — М.: Альпина нон-фикшн, 2019. — 358 с. — ISBN 978-5-91671-893-5.
- Нэйш Д.[англ.], Барретт П. Динозавры. 150 000 000 лет господства на Земле / науч. ред. А. О. Аверьянов, д-р биол. наук. — М.: Альпина нон-фикшн, 2019. — 223 с. — ISBN 978-5-91671-940-6.
- Татаринов Л. П. Ящеротазовые динозавры, или заврисхии // Очерки по эволюции рептилий. Архозавры и зверообразные. — М. : ГЕОС, 2009. — С. 68—125. — 377 с. : ил. — (Труды ПИН РАН ; т. 291). — 600 экз. — ISBN 978-5-89118-461-9.
На английском языке
править- Bakker R. T. The Dinosaur Heresies: New Theories Unlocking the Mystery of the Dinosaurs and Their Extinction (англ.). — New York: William Morrow and Company, 1986. — 481 p. — ISBN 978-0-688-04287-5.
- Benton M. J.[англ.]. Vertebrate palaeontology[англ.] (англ.). — 4th ed. — Wiley-Blackwell, 2015. — 480 p. — ISBN 978-1-118-40755-4. — ISBN 978-1-118-40684-7.
- The Complete Dinosaur (англ.) / M. K. Brett-Surman, Holtz T. R. Jr., J. O. Farlow, eds. — Bloomington, IN: Indiana University Press, 2012. — 1272 p. — (Life of the Past). — ISBN 978-0-253-00849-7.
- Brusatte S. L. Dinosaur Paleobiology (англ.) / ed. by M. J. Benton[англ.]. — University of California Press, 2012. — 336 p. — ISBN 978-0-520-25408-4.
- Molina-Pérez R., Larramendi A. Dinosaur Facts and Figures: The Theropods and Other Dinosauriformes (англ.) / Illustrations by A. Atuchin and S. Mazzei. — Princeton, New Jersey: Princeton University Press, 2019. — 288 p. — ISBN 978-0-691-18031-1.
- Molina-Pérez R., Larramendi A. Dinosaur Facts and Figures: The Sauropods and Other Sauropodomorphs (англ.) / Illustrations by A. Atuchin and S. Mazzei. — Princeton, New Jersey: Princeton University Press, 2020. — 272 p. — ISBN 978-0-691-20297-6.
- Owen R. Palaeontology, or a Systematic Summary of Extinct Animals and their Geological Relations (англ.). — Second Edition. — Edinburgh: A & C Black, 1860. — 463 p.
- Paul G. S.[англ.]. Predatory Dinosaurs of the World: A Complete Illustrated Guide (англ.). — New York: Simon & Schuster, 1988. — 464 p. — ISBN 978-0-671-61946-6.
- Paul G. S.[англ.]. The Princeton Field Guide to Dinosaurs[англ.] (англ.). — Princeton: Princeton University Press, 2016. — 360 p. — ISBN 978-0-691-16766-4.
- The Dinosauria / D. B. Weishampel, P. Dodson, H. Osmolska, eds. — 2nd ed. — Berkeley: University of California Press, 2004. — 861 p. — ISBN 0-520-24209-2.
- Phylonyms: A Companion to the PhyloCode (англ.) / de Queiroz K.[англ.], Cantino P. D., Gauthier J. A., eds. — Boca Raton: Taylor & Francis Group, CRC Press, 2020. — 1352 p. — ISBN 978-1-138-33293-5.